LABORATORIO DE LÍPIDOS Y EXOCITOSIS ACROSOMAL
Responsable: Dra. Silvia Belmonte
El espermatozoide humano es una célula terminal, muy polarizada cuya única función es transferir el material genético al ovocito. Para que esto ocurra debe secretar el contenido de su gránulo apical denominado acrosoma. Este proceso se denomina exocitosis o reacción acrosomal, tiene un mecanismo complejo que involucra la apertura de múltiples poros de fusión y está altamente regulado.
La fusión de membranas en todos los modelos conocidos utiliza una serie de proteínas altamente conservadas. Nuestro laboratorio enfoca la exocitosis desde el punto de vista lipídico. El principal objetivo es develar cómo los diferentes lípidos que constituyen las membranas participan activamente en el proceso de fusión cumpliendo un rol específico en la exocitosis. Los lípidos tienen funciones estructurales, son moléculas que intervienen en numerosos mecanismos de transducción de señales, actúan como plataformas de unión para diferentes proteínas y afectan la curvatura de membrana.
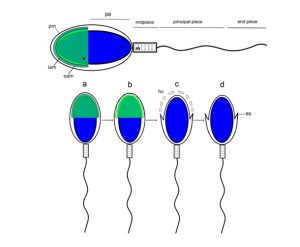
El esquema superior muestra características del espermatozoide humano. El flagelo se divide en tres regiones, en la cabeza se ubica el núcleo y el acrosoma, En el esquema inferior se detallan los cambios morfológicos que ocurren en la cabeza del espermatozoide humano durante la exocitosis acrosomal (Suhaiman el al, Methods Mol Biol, 2021).
Líneas de investigación
I- Mecanismos moleculares activados por distintos esfingolípidos en la exocitosis acrosomal
Hemos develado el complejo mecanismo de transducción de señales modulado por esfingosina 1-fosfato, ceramida y ceramida 1-fosfato. Pretendemos determinar la conexión de estas vías con inductores fisiológicos de la reacción acrosomal (zona pellucida ovocitaria y progesterona).
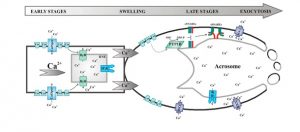
Resumen gráfico de la secuencia de señalización de eventos que conectan la ceramida con la movilización interna de calcio y señales de calcio externas durante la secreción (Vaquer, Suhaiman et al, 2020).
II- Correlación entre composición de fosfolípidos y la curvatura de membrana en la exocitosis acrosomal
Diacilglicerol (DAG) promueve la exocitosis acrosomal en espermatozoides humanos permeabilizados induciendo un ciclo de retroalimentación positiva, dependiente de la activación de PKC y PLD1, que provee fosfatidilinositol 4,5-bifosfato (PIP2) continuamente durante el proceso exocítico. El PIP2 producido es necesario para incrementar la síntesis de inositol-1,4,5-trifosfato (IP3) y promover la salida de calcio del reservorio acrosomal. ARF6 regula el recambio fosfolípidico durante la reacción acrosomal, particularmente las concentraciones de PIP2 y ácido fosfatídico. Ambos lípidos son necesarios para la exocitosis y afectan la curvatura de membrana durante el proceso.
III- Moléculas que regulan la composición de fosfolípidos durante la exocitosis.
En este sentido estamos estudiando la posible activación de PLCepsilon directamente por DAG. Postulamos que este lípido activa un GEF (factor intercambiador de nucleótidos, CalDAG-GEF) para la GTPasa Rap1. Rap-GTP activaría PLCe (Figura 3). Hemos determinado previamente que la síntesis de PIP2 ocurre durante la exocitosis acrosomal, consideramos de importancia determinar si alguna isoforma de la quinasa que lo produce está presente en la gameta masculina. Pretendemos establecer la presencia y función de la PI4P5K en el espermatozoide humano (Figura 3).
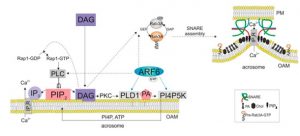
Modelo donde se muestra la importancia de los lípidos como moléculas de señalización y en la curvatura de membrana durante la exocitosis acrosomal.